马上注册,结交更多好友,享用更多功能,让你轻松玩转社区。
您需要 登录 才可以下载或查看,没有账号?立即注册

x
目前在家禽生产中,热应激带来的危害已经成为一个普遍的问题!
热应激是综合环境因素所导致的结果,包括气温、湿度和风速等,其中温度起着重要的作用 。 当环境温度超过商品肉鸡最佳生长温度时,就可能引发热应激反应,导致体温升高 、呼吸频率加快 、采食量和体增重下降 、胸肌肉产量降低 和肉品质下降等。
Lin等总结了几种缓解热应激的常见策略,包括基因策略、营养策略、饲养策略和环境策略。 其中为鸡舍提供良好的通风环境,在肉鸡生产过程中具有十分重要的意义,这一点在热带和亚热带地区显得尤为突出。最早开始研究通风对于家禽的影响要追溯到20世纪 60年代。 Drury 等首次提出了“ 肉鸡生产得益于风速产生的风冷作用” ,并得出风速可以改善肉鸡生产性能。 随后的几十年,美国、以色列和巴西等国先后开展了相关研究。 在20世纪末,通风管理已经在家禽生产中得到广泛的应用。 虽然夏季通风能够缓解热应激和降低死亡率,但是能够系统阐释的数据少之又少,机理尚不完全清楚。
据统计,国外专项研究风速对家禽影响的实验团队屈指可数,发表的论文也不过 20余篇,国内仅有陶秀萍较早对温湿风进行了初步的研究,并得出了肉鸡温湿风指数模型,探究了风速对肉鸡生理生化的影响。因此,本文将扼要总结家禽热平衡及其调节,风速对可感散热和潜热散热的贡献,并结合对生产性能的陈述,阐释风速在家禽热平衡调节中的作用。
1 家禽热平衡及其调节
1.1 家禽的热平衡
所谓热平衡,即产热和散热达到一种动态平衡的状态 。 家禽热平衡的维持需要机体时刻进行相应的调节,主要分为物理性调节和化学性调节。 其中物理性调节包括生理和行为 2 方面。 肉鸡处在高温或低温环境中,首先会引起皮肤血管的舒张或收缩,进而增加或降低皮肤血流量和皮肤温度,增加或减少与外界的温差,以利于热平衡的维持。
家禽行为在热平衡的维持中起着十分重要的作用。 而化学性调节主要是在物理性调节不能保持热平衡时,依靠调整体内代谢率来增加或减少产热量。家禽体内存在一套完整的机制(图 1) ,用于维持热平衡。 温度、湿度和风速综合表征环境的热负荷量,作用于家禽体热感受器 ,包括外周温度感受器和中枢温度感受器。 这些器官将感受的热信息上传至中央处理器,调节体温的中枢结构存在于从脊髓到大脑皮层的整个神经系统内,但是体温调节的基本中枢位于下丘脑。 其中视前区下丘脑前部(PO/AH )是体温调节中枢的关键部位 。 热信息经过中央处理器处理后,通过神经或 / 和内分泌途径支配效应器产生反应 ,调节散热量,完成热平衡的调节,以达到机体热平衡的状态。
图 1 家禽感受环境刺激和热平衡调节反应示意图
1.2 家禽散热方式及其分配模式
家禽的散热方式分 2 类:可感散热( SHL ) 和潜热散热 ( LHL)。SHL 又称为 “ 非蒸发散热”或 “ 显热散热” ,包括辐射、传导和对流,皆为物理散热方式。
LHL 又可称为“不可感散热”或“蒸发散热” ,包括皮肤和呼吸道水分的蒸发。 由于家禽身体大都被羽毛覆盖,没有汗腺,无法靠体表直接出汗散热;同时传导需要温度不同的介质相互接触,对于家禽的散热贡献不大。
实际上家禽的散热方式包括以下2 种:
1 )以辐射和对流为主的SHL ;
2) 以呼吸蒸发机制和皮 肤蒸发机制为主的 LHL。
家禽体表面和外界的温度差是 SHL 的驱动力,只要温度差是正的,就会一直在进行,而且全程是物理作用,不会消耗额外的能量。 以热喘为重要散热方式的 LHL 则恰恰相反,这个过程需要消耗大量的能量。不过,随着环境温度的上升,SHL 所占的散热比重逐渐降低。 低温和适温情况下(13 ~ 25 ℃ )肉鸡主要通过辐射和对流进行散热;当温度上升至30 ℃时,LHL 所占的比重逐渐升高;而当外界温度达到 35 ℃时,LHL 量占总散热量的 90%。 肉鸡在不同环境温度的散热分配见表 1。
2 风速对家禽热平衡调节的影响
从为数不多的研究中可以发现,风速主要通过对家禽能量平衡、水平衡的调节进而影响 SHL、LHL 量及其比例来调节热平衡。
2.1 风速对 SHL 的作用
总结风速对 SHL 作用的研究,以2004年 Ya?hav 等运用红外热成像技术研究 SHL 为标志可分为 2 个阶段。第 1 阶段( 2004 年以前) :在这一阶段,研究者基本认为 SHL 在高温应激时对热调节的作用不大。风速的大小影响对流散热,进而影响 SHL。风速增大,任何热物体的对流传递的热量也随之增大 。 其实对流散热现象是个复杂的过程,可以用“ 热边界” 来解释。通过对火鸡的研究发现,高于温度适中区的偏热情况下,SHL 量会随着风速的升高而增加,但是在高温情况下 SHL 的占比会降低。SHL 量同样受到日龄的影响。 Jurkschat 等经过试验得出了一套公式阿—用于描述火鸡日龄、风速及环境温度跟 SHL 量的相互关系。
研究同样发现,风速的增加可以升高 SHL 量。 Mitchell 等使用“风道热量计” 监测风速对 15周龄来航鸡SHL 量的影响。 结果表明,在 30 ℃ 高温情况下,风速从 00.2 m /s 上升到 1.2 m/s,SHL 量从 7.6W升高 到 12.6 W。 Timmons 等设置不同温度(35、38、41 ℃ )和风速( 0.25/1.0/2.0m/s)组合,研究成年来航鸡的散热量的变化。 结果发现,在35 ℃高温情况下,增加风速能够使得更多的热量通过对流的方式散发。
尽管风速可以提高 SHL量,Hillman等 却总结前人研究后认为,当环境温度高于热中性区上限时,SHL 对家禽散热的作用不大。 因为对于全羽鸡来讲,身体被羽区体表与环境之间的温差很小,并且裸露的部位十分有限,仅包括腿部、头部、肉垂和鸡冠,所以 Tzschentke 等认为高温下SHL 在体热调节中的作用可以被忽略不计。第2 阶段(2004年以后) :发现 SHL 在高温环境下肉鸡能量平衡调节中起主要作用。这得益于红外热成像技术的运用。 在这之前红外热成像技术就已经成功地应用到哺乳动物和鸟类的表皮温度的测量上 。 而且红外热成像辐射计测量得出的表皮温度要比传统的热电偶更加精确 ,同时也能准确测量拍摄区域的面积。 Yahav 等运用红外热成像技术测定了肉鸡各个部位的对流散热和辐射散热,研究了风速对于 SHL 量的贡献和SHL 量占维持能的百分比,得出,在35 ℃ 、60%相对湿度情况下,0.8、1.5、2.0和 3.0 m /s 4 种不同的风速下肉鸡 SHL 量占到总维持能的比例分别24.1%、29.1%、36.8%和44.7%,发现 SHL 量最高可以占到维持能的近 45%,据此认为风速可提高SHL 量,其在肉鸡能量调节中起主要作用。
2.2 风速对 LHL 的作用
LHL 通过影响家禽体内水代谢调节。 一部分潜热通过家禽皮肤散发( 隐形蒸发和非显性出汗) ,而其余大部分潜热通过呼吸蒸发散失。 环境温度过高,家禽呼吸频率急剧增加,热量通过呼吸蒸发水分散失,严重时导致水平衡和酸碱平衡失调。 Belay 等证明,相比于 24 ℃ ,暴露在35℃4 h 下的肉鸡失水量升高了 64%。Teeter 等 发现,4 周龄肉鸡热应激( 32 ~ 41 ℃ ) 导致呼吸率显著升高,水分流失、二氧化碳分压下降、碳酸根离子( HCO- 3 )浓度下降、血液 pH 显著升高,出现呼吸性碱中毒。Simmons 等研究发现, 环境温度在29 ~35℃之间时,随着风速增加(从61m /in 增加到183 m /min),肉鸡LHL量 下 降。 Yahav 等将6 ~ 7 周龄肉鸡饲养于35 ℃ (高温) 、相对湿度 60%的环境中,研究不同风速对水平衡的影响时发现,1.5 和 2.0 m /s 风速下体温、精氨酸催产素浓度和血浆渗透压都显著低于低风速(0.8m/s) 和高风速(3.0m /s)下;失水状态下,精氨酸催产素浓度和血浆渗透压都会升高。 这表明,较低风速(0.8m/s)时,肉鸡热量大部分还是依靠热喘散发,高频率的呼吸会过多带走体内的水分,导致水平衡的破坏;而高风速(3.0m/s) 情况下,一方面机体内水分大量通过皮肤散发,不利于水平衡的维 持。 可见,适宜风速通过调节水平衡减少LHL 量。
2.3 对散热分配方式的影响
风速影响家禽的散热方式。 Simmons 等研究了风速对 SHL 和 LHL 量的影响,发现环境温度在 29 ~ 35 ℃之间时,总的散热量不会随着风速的增加(从 61 m / min 增加到 183 m / min)而改变,相反,却处于一个相对稳定的范围;风速上升,SHL量增加,LHL 量下降;而温度上升,却伴随着 SHL量下降,LHL 量上升;而且,当风速为2.0m / s左右时,无论温度如何上升,总散热量会保持相对恒定;风速提高,LHL 转化为 SHL,能够促进肉鸡的生产性能。简而言之,以红外热成像技术研究SHL为节点,发现SHL在高温环境下肉鸡能量平衡调节中起到主要作用,证实了风速对 SHL 的重要影响;同时发现,适宜风速通过调节水平衡减少 LHL量;高温下风速的提高促使 LHL 向 SHL 转化,进而增加SHL 的占比,相应减少 LHL 的占比,提高能量的利用率。 总之,风速通过对家禽能量平衡、水平衡的调节,进一步影响 SHL、LHL 及其比例,从而达到调节热平衡的目的。
3 风速对于家禽生产性能的作用及影响因素
自从 Drury 等[8-9]首次发现“肉鸡生产得益于风速产生的风冷作用”以后,许多试验证实了风速调节对家禽生产性 能的提高作用 。生产性能的提高是风速对高温环境下家禽热平衡调节的结果。风速对生产性能是否提高取决于风速大小、家禽日龄、环境温度、饲养方式和通风方式等因素。
Mitchell研究表明,在高温、高湿极端环境下,提高风速可能是冷却降温的唯一切实可行的方法。 Ruzal 等[40]研究风速在高温下对蛋鸡的影响时发现,35 ℃ 情况下,3.0 m / s 显著提高鸡蛋产量,而低风(0.5m/s) 不利于鸡蛋的产量和品质。据 Lott等报道, 环境温度为28 ℃ 时,125 m / min的风速较 15m/smin( 当风速小 于0.25m / s时被认为是无风静止状态,可忽略其影响)能够提高 4 ~ 6 周龄肉鸡的体增重和饲料转化率。 Yahav 等和 Ruzal等研究了不同的风速(试验 1:0.5、1.5、2.0和 3.0m / s;试验 2:1.0、1.5、2.0和 2.5m / s) 对在高温( 35 ℃ ) 、相对湿度60%环境下单笼饲养肉鸡、火鸡生产性能的影响。结果表明,2 次试验中暴露在1.5 和2.0 m/s 风速下 4 ~ 7 周龄的肉鸡体增重、采食量和饲料转化率显著升高。 低风速(0.5和 1.0 m / s)组的生产性能要低于高风速(2.5 和 3.0m / s) 组,并得出最佳风速范围是在 1.5 ~ 2.0 m / s。 同时 Yahav 等提出风速影响生产性能过程中存在一个温度“拐点” 。在环境温度低于 30 ℃ 时,通风增加维持能耗,降低生长效率。在一种日循环模式温度( 22—32—22 ℃ ) 中,May等的研究表明,2 m / s 的风速会降低 21~49日龄肉鸡的饮水量,提高采食量、体增重和肉重比。 但是 Simmons 研究了另一种循环模式温度(25—30—25 ℃ ) ,露点温度为 23 ℃ 时,21 ~28 日龄群养的罗斯肉公鸡在不同风速下(3、2 和<0.25m/s)的体增重并没有显著的影响。 然而,随着肉鸡的生长,风速的作用逐渐凸显。 到 42 ~ 49日龄时,3m /s 风速下的体增重和料重比要显著高于 2m/s 和<0.25m/s 组。 Dozier 等 得出了相似的结果。 在 25—35—25 ℃ 循环模式温度,露点温度为 23 ℃ 时,21 ~ 49 日龄的肉公鸡处在 3 m/s高风速下的生产性能要高于 2 m/s 相对低风速状态下。Dozier 等研究了通风方式对日循环高温(25—35—25℃ )下肉鸡生产性能的影响,试验结果表明,昼夜24h 高风速(2.79m/s)组 37 ~ 51 日龄肉公鸡的体增重和饲料转化率明显高于仅白天12 h 高风速组肉鸡,但采食量和死亡率没有显著差异。综合以上试验结果可以发现,风速对家禽生产性能的作用与环境温度、风速大小、饲养方式(单养和群养) 、日龄和通风方式有关。
4 小 结
本文通过总结家禽热平衡及其调节,风速对SHL 和 LHL 的作用,并结合对生产性能的陈述,阐释了风速在家禽热平衡调节中的要作用。 风速增加高温环境下家禽 SHL 量,适宜风速时的SHL在能量平衡调节中起主要作用;通过维护水平衡减少 LHL 量;通过增加 SHL 占比,相应减少LHL 占比,提高能量的利用率,从而提高体增重和饲料转化率。 风速对生产性能的提高作用取决于风速大小、肉鸡日龄、环境温度、饲养方式和通风方式等。 今后应进一步研究风速在不同高温、湿度下家禽能量平衡和水平衡调节中的作用及其途径,系统阐释风速调节热平衡的作用机制,为合理通风提供科学依据。
作者:张少帅 张敏红
| 













 发表于 2021-7-1 10:14:28
发表于 2021-7-1 10:14:28


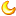

 发表于 2021-7-1 13:05:43
发表于 2021-7-1 13:05:43
